Haben Sie Vorsorge getroffen?
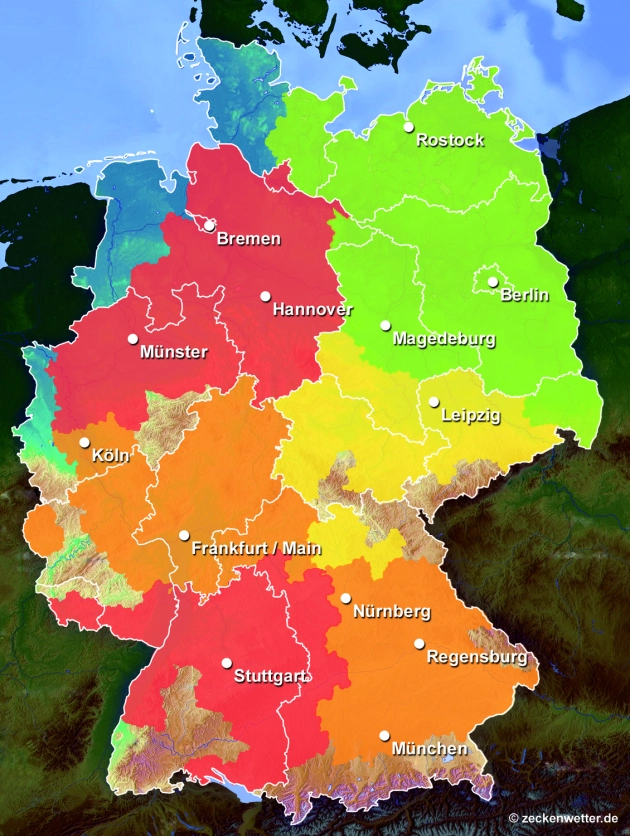
Schauen Sie am besten direkt unter zeckenwetter.de nach, wie hoch die Gefahr von Zeckenbissen in ihrer Gegend zurzeit ist. Hier finden umfangreiche Informationen auch für Tiere. Beugen Sie auch mit einer passenden Impfung vor.
Über die Ausbreitung der Zecken nach Norden
Information zu den aktuellen Gefahren für die Gesundheit. Von Olaf Kahl & Hans Dautel.
Die Auwaldzecke breitet sich aus
Eine Vektorzecke überträgt Krankheiten vor allem auf Tiere. Hans Dautel und Olaf Kahl.